

Durante 2017 estamos celebrando el 50 aniversario de la publicación en la revista PNAS del primer experimento de evolución in vitro de ácidos nucleicos, por el grupo de Sol Spiegelman. Aquel logro científico resultó fundamental en la investigación sobre el origen de la vida, y abrió la puerta a la obtención de novedosas herramientas biotecnológicas. Además, supuso el nacimiento de un monstruo…
El año 1953 suele considerarse el punto de partida de la biología molecular, debido al descubrimiento de la estructura en doble hélice del DNA y a la primera determinación de la secuencia de aminoácidos de una proteína, la insulina bovina. Ese mismo año, el famoso experimento realizado por Stanley L. Miller inauguró la química prebiótica como disciplina experimental, al demostrar que los aminoácidos y otras biomoléculas pueden producirse como resultado de reacciones químicas a partir de precursores moleculares muy sencillos. Gracias a todo ello, la década de 1960 se inició con la sensación de que la química orgánica y la bioquímica acabarían explicando los procesos fundamentales relacionados con el origen y la evolución de la vida.
Sin embargo, en la comunidad científica de la época se asumía que los procesos evolutivos operaban únicamente sobre los organismos (unicelulares o pluricelulares) capaces de reproducirse autónomamente, así como sobre los virus, cuya replicación se realiza a expensas de la maquinaria metabólica de las células a las que parasitan. Pero un investigador brillante, el biólogo molecular norteamericano Sol Spiegelman, se planteó a mediados de esa década algo que nadie había abordado experimentalmente antes: ¿podría la evolución regir el comportamiento de conjuntos de moléculas (en concreto, ácidos nucleicos) sometidos a amplificación y selección in vitro, es decir, en un tubo de ensayo y sin células ni virus en el medio? Dicho de otra forma: ¿puede haber evolución por selección natural en ausencia de seres vivos y de entidades biológicas sub-celulares como los virus?

Replicantes en el laboratorio
Sol Spiegelman había nacido en Nueva York en 1914, y tras estudiar en la Universidad de Columbia (Nueva York) y en la Universidad Washington (San Luis, Missouri) se incorporó como Profesor de Microbiología a la Universidad de Illinois (Urbana, Illinois) en 1949. A partir de 1963, entre otros sistemas experimentales comenzó a trabajar en su laboratorio con la molécula de ácido ribonucleico (RNA) que constituye el genoma del bacteriófago Qβ, un virus que infecta a la bacteria Escherichia coli. Este “fago”, de morfología esférica y simetría icosaédrica, es uno de los virus más pequeños que se conocen ya que el diámetro de su cápsida es de tan solo 25 nanómetros (unas cuarenta veces menor que su célula hospedadora). El material genético viral es una cadena sencilla de RNA con “polaridad positiva” [RNA-Qβ(+)], es decir, cuando se libera en la célula infectada puede ser traducido directamente por los ribosomas de E. coli, produciéndose así las proteínas del fago.
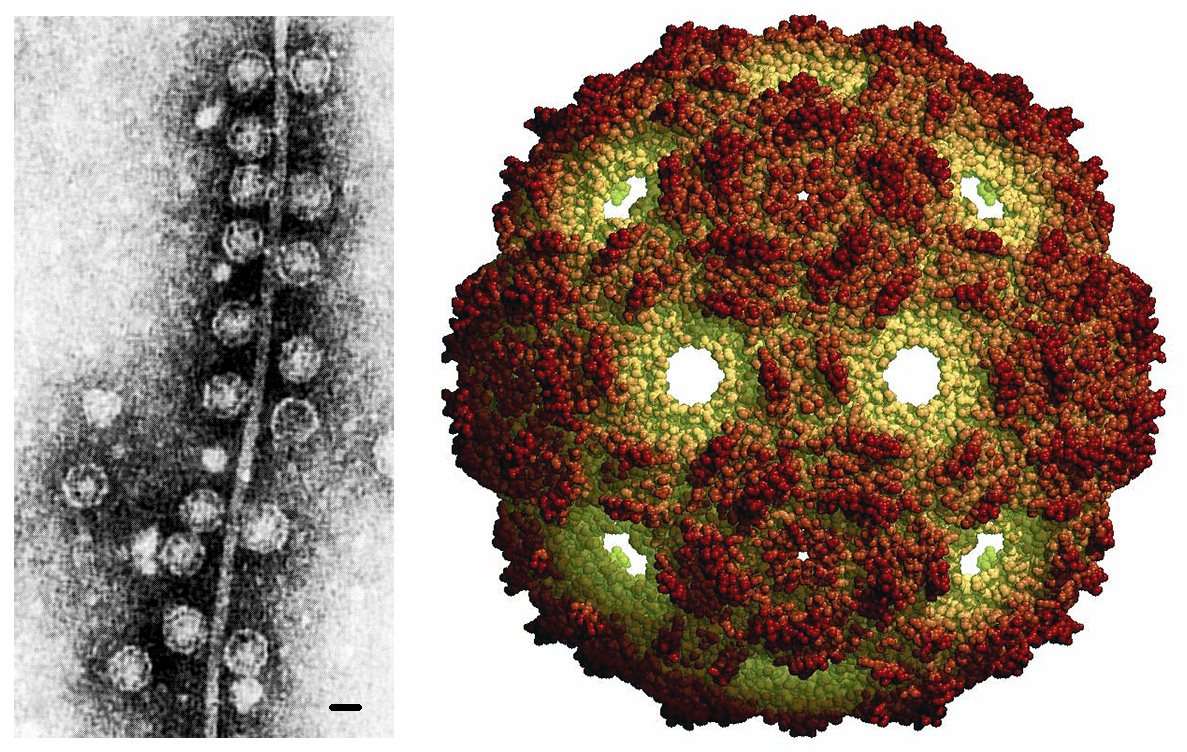
Hoy sabemos que la longitud del genoma del fago Qβ es de tan solo 4.220 nucleótidos (unas mil veces menor que el de su hospedador, pues el DNA genómico de E. coli tiene 4,6 millones de pares de nucleótidos), y en él están codificadas únicamente cuatro proteínas. Entre ellas, la Qβ replicasa resultaba clave para el experimento planteado ya que es la enzima encargada de la replicación del genoma viral, un proceso que se realiza en dos pasos [RNA-Qβ(+) → RNA-Qβ(-) → RNA-Qβ(+)]. Y ahí radicaba una característica fundamental: como el propio Spiegelman demostró en 1965 (Habuna y Spiegelman, 1965), a pesar de que en el interior de una E. coli infectada se requería que esta enzima viral interaccionara con tres proteínas celulares para replicar el genoma del bacteriófago, la Qβ replicasa era capaz por sí misma de copiar el material genético del virus cuando éste se colocaba en un tubo de ensayo junto a los reactivos químicos necesarios.
A partir de ahí, el siguiente experimento que se llevó a cabo en el grupo de Spiegelman fue muy imaginativo, aunque relativamente sencillo desde el punto de vista experimental: en un tubo de ensayo con 250 microlitros de una disolución acuosa (conteniendo las sales y el estabilizador de pH requeridos en el proceso) se puso un número pre-calculado de copias del RNA genómico completo del fago Qβ junto a la cantidad suficiente de la proteína Qβ replicasa, ambos previamente extraídos del virus y debidamente purificados. Como la replicación del genoma se iba a realizar fuera del entorno celular, ese medio debía suplementarse con los ribonucleótidos (monómeros de la molécula de RNA: ATP, CTP, GTP y UTP) necesarios para copiar el genoma molde.

En tales condiciones y a una temperatura de trabajo de 35°C, la Qβ replicasa hacía copias del RNA genómico viral, y cuando los ribonucleótidos se agotaban era preciso transferir una fracción (en sus experimentos, 20 microlitros) de la mezcla de reacción a otro tubo de ensayo con sustratos frescos. Durante este proceso, que implicó 15 pases seriados, se utilizó el sistema experimental más preciso de la época para seguir la producción de RNA.
Como resultado, Spiegelman publicó el 1 de septiembre de 1965 un artículo en la prestigiosa revista Proceedings of the National Academy of Sciences of the USA (PNAS) en el que mostraba cómo era posible sintetizar in vitro un ácido nucleico con capacidad para propagarse en pases seriados, manteniendo su infectividad en E. coli (Spiegelman y col., 1965). Este trabajo se hizo popular rápidamente y las crónicas de la época lo describieron como “el experimento soñado” en biología, ya que aparentemente permitía producir una entidad auto-replicativa en el tubo de ensayo.
De hecho, durante los días posteriores a la publicación del artículo, ciertas entrevistas que hicieron a Spiegelman reflejaron un sesgo que sigue siendo habitual hoy en día: mientras el científico decía que “por primera vez se ha puesto a punto un sistema que permite el análisis de las bases moleculares que subyacen a la replicación de un ácido nucleico con capacidad para auto-propagarse”, algunos periodistas desinformados (o convencidos de que la realidad no debe estropear un buen titular) resumieron el trabajo como “Se ha creado vida en el tubo de ensayo”. Ante tales expectativas, Spiegelman pronunció una conferencia en su universidad el día 30 de septiembre… y la afluencia de alumnos y profesores fue tal que se llenaron rápidamente los más de 350 asientos del salón de actos, teniendo que intervenir la policía para liberar los abarrotados pasillos y mantener el orden.

¿Y si el RNA pudiera evolucionar in vitro?
Una vez puesto a punto con éxito el sistema experimental, Spiegelman se propuso comprobar si era posible recrear procesos evolutivos empleando pases seriados tubo a tubo. Aquella fue una época complicada porque, debido a la fama cosechada por su laboratorio, tenía que atender a decenas de científicos visitantes (fundamentalmente enzimólogos) y alumnos que deseaban trabajar en él. Además, su secretaria estuvo hospitalizada con neumonía durante varias semanas, y la situación económica tampoco resultaba fácil ya que la Guerra de Vietnam había obligado a reducir la financiación de su universidad.
Pero todo esto no alteró la senda científica que Spiegelman se había trazado. Dado que en sus experimentos in vitro la única variable sometida a presión selectiva era la velocidad de copia del RNA, diseñó un sistema de selección para favorecer a las moléculas de RNA molde más rápidas en ser replicadas, que presumiblemente serían las más cortas. Con ello dispondría, en un mismo sistema experimental, de los dos ingredientes clave de la evolución: replicación y selección. Así, el tiempo de incubación de la reacción fue reduciéndose según el siguiente esquema: 20 minutos (pases 1 al 13), 15 min (14-29), 10 min (30-38), 7 min (39-52) y 5 min (53-74). Los resultados del experimento, aún más convincentes de lo esperado, fueron enviados por Spiegelman a la revista PNAS el 18 de mayo de 1967 y se publicaron en el número del 1 de julio de ese año (Mills y col., 1967).

Algo que mostraron los resultados de este revolucionario trabajo fue que tras solo 4 pases dejaban de aparecer las secuencias genómicas “biológicamente competentes”, es decir, aquellas suficientemente completas como para mantener su capacidad de infectar a E. coli. De hecho, a partir del pase 9 ya no se detectaban moléculas de RNA con la longitud del genoma viral original. A lo largo de todo el resto del proceso se observó que el RNA molde seguía acortándose y en paralelo su velocidad de replicación iba siendo cada vez mayor. Al terminar los 74 pases previstos, la molécula resultante tenía (según se calculó posteriormente) sólo 550 nucleótidos y era copiada por la Qβ replicasa a una velocidad 15 veces mayor que el genoma original. No cabía ninguna duda: el RNA realmente había evolucionado dentro del tubo de ensayo.

El nacimiento de un monstruo
Los resultados del artículo de 1967 tuvieron, además del lógico reconocimiento científico, una insospechada repercusión en la prensa de la época. Así, esa molécula de RNA obtenida (inexistente como tal en la naturaleza) comenzó a denominarse popularmente “Monstruo de Spiegelman”, ante la sospecha de que un ser vivo artificial y potencialmente peligroso hubiera sido fabricado en un laboratorio de Illinois. A ese temor pudo contribuir involuntariamente el propio Spiegelman, ya que en sus artículos llamaba al RNA resultante del proceso “entidad replicativa” o incluso “ácido nucleico auto-replicativo”. La segunda denominación resultaba impactante pero en esencia es incorrecta, ya que tal RNA no era autónomo: para replicarse requería la enzima Qβ replicasa, además de un aporte constante de monómeros.
Desde el punto de vista de la bioseguridad, ese pequeño “monstruo” ni siquiera era capaz de infectar células de E. coli… y a temperatura ambiente tardaba muy poco en degradarse por las enzimas presentes en cualquier medio. Pero en el imaginario colectivo despertaba los mismos miedos infundados que años después generaría la llegada de la ingeniería genética en bacterias, la clonación, las plantas y animales transgénicos o la biología sintética.
En cualquier caso, lo fundamental de este experimento es que mostró cómo los mecanismos evolutivos no afectan únicamente a los organismos celulares y a los virus, sino que también rigen el comportamiento de conjuntos de moléculas de ácidos nucleicos sometidas a procesos de amplificación y selección in vitro. De esta forma quedaba inaugurada una disciplina científica denominada “evolución experimental in vitro”, también llamada en ocasiones “evolución darwiniana extracelular” o, simplemente, “evolución molecular”.
La continuación del trabajo de 1967 se centró en analizar detalladamente los resultados obtenidos, gracias a que en esa época comenzaban a desarrollarse las técnicas de secuenciación de RNA. En paralelo, la modificación de las presiones selectivas aplicadas permitió obtener diferentes variantes genéticas de los RNAs resultantes del proceso evolutivo in vitro. Además, mediante experimentos publicados cinco años después, el grupo de Spiegelman logró seleccionar un “monstruo” aún más reducido que el original, con tan solo 218 nucleótidos (Kacian y col., 1972), que pudo ser secuenciado un año después (Mills y col., 1973).

En ese RNA tan corto se habían eliminado todas las secuencias innecesarias para la replicación de la molécula in vitro, hasta seleccionarse únicamente una región directamente implicada en la unión a la replicasa. La molécula ganadora de la carrera evolutiva, veinte veces más corta que el genoma viral del que derivaba, abría la puerta a interesantes avances en ciencia básica (relacionados con el estudio del propio proceso de replicación de los ácidos nucleicos, o con el origen de la vida y la evolución precelular) así como en virología (al poder diseñarse moléculas con capacidad de interferir en la replicación de ese mismo virus, e incluso de otros con relevancia clínica).
A pesar de la trascendencia de sus descubrimientos, Spiegelman afirmaba humildemente que cualquier otro investigador podría haber dado ese mismo paso, ya que todos los avances posibles en una época determinada acaban por producirse. Así, según comentaba en una entrevista que le realizaron en 1975: «Los científicos son los catalizadores de lo inevitable. Yo llamo a esto mi principio de ‘dispensabilidad’.»

Tras estos trabajos pioneros, Sol Spiegelman continuó siendo un referente mundial en biología molecular, realizando aportaciones muy relevantes en el campo de la ingeniería genética hasta su muerte en 1983. De hecho, en octubre de este año se va a celebrar en la que fue su universidad un simposio para recordar su legado, en el que participarán varios de los biólogos moleculares norteamericanos actuales más relevantes. Entre ellos se encuentran Jack W. Szostak, Larry Gold y Gerald F. Joyce, protagonistas de la nueva era de la evolución molecular in vitro iniciada en 1990 y (como comentaremos en otra ocasión) cuyos hallazgos siguen resultando fundamentales en investigación básica y aplicada.
Darwin en el tubo de ensayo
Cincuenta años después del primer experimento de evolución in vitro, quienes trabajamos en temas relacionados con el origen y la evolución de la vida o utilizamos herramientas biotecnológicas basadas en ácidos nucleicos estamos en deuda con el grupo de aquel investigador visionario y tenaz llamado Sol Spiegelman.
También deberían recordar sus logros el resto de científicos, y todos los que utilizan el pensamiento racional como forma de explicar los fenómenos que ocurren en la naturaleza. La obra de Charles R. Darwin y Alfred R. Wallace a mediados del siglo XIX, así como los avances en paleontología, biología molecular, filogenia y genómica comparada durante el último siglo, han demostrado que la evolución no es un sistema de creencias ni una teoría debatible, sino una realidad que ocurre constantemente en todos los rincones de la biosfera, que nunca ha podido ser falsada científicamente, y que es consustancial al propio fenómeno vital. Así, como reconocía el título de un famoso ensayo publicado en 1973 por el genetista Theodosius Dobzhansky: “Nada en Biología tiene sentido si no es a la luz de la evolución”.
Pero además de explicar la realidad biológica, como hemos recordado en estas líneas el trabajo de Spiegelman demostró que la evolución puede comprobarse en pocas horas en un laboratorio convencional de biología molecular, dentro de un tubo de ensayo… y ante la mirada de cualquier observador, independientemente de lo que éste crea o de si preferiría que la naturaleza hubiera sido diseñada por algún ser imaginario.
Referencias:
Haruna I, Spiegelman S (1965). Specific template requirements of RNA replicases. Proc. Natl. Acad. Sci. USA 54: 579-587.
Spiegelman S, Haruna I, Holland IB, Beaudreau G, Mills DR (1965). The synthesis of a self-propagating and infectious nucleic acid with a purified enzyme. Proc. Natl. Acad. Sci. USA 54: 919-927.
Mills DR, Peterson RL Spiegelman S (1967). An extracellular Darwinian experiment with a self-duplicating nucleic acid molecule. Proc. Natl. Acad. Sci. USA 58: 217-224.
Kacian DL, Mills DR, Kramer FR, Spiegelman S (1972). A replicating RNA molecule suitable for a detailed analysis of extracellular evolution and replication. Proc. Natl. Acad. Sci. USA 69: 3038-3042.
Mills DR, Kramer FR, Spiegelman S (1973). Complete nucleotide sequence of a replicating RNA molecule. Science 180: 916-27.
Doctor en Ciencias Químicas (esp. Bioquímica y Biología Molecular) por la Universidad Autónoma de Madrid. Es Científico Titular del CSIC en el Dpto. de Evolución Molecular del Centro de Astrobiología (CSIC-INTA, asociado al NASA Astrobiology Institute), donde dirige un grupo que trabaja en los campos del origen de la vida, los virus con genoma de RNA y la bionanotecnología. Cuando su investigación le deja tiempo se entrega con pasión a la divulgación científica. Y desde que tiene (o no) uso de razón escribe poesía y relatos. Resultado: curiosidad por casi todo, muchos libros leídos, algunos escritos… y confianza en que la Tercera Cultura puede mejorar el mundo.