

Después de un largo invierno y una triste primavera, ya está aquí el período estival y, como todos los años, empezamos a recibir las primeras noticias de incendios forestales que azotan nuestros bosques y destruyen nuestro entorno natural. El año pasado 2019 se contabilizaron alrededor de 10,883 incendios incluyendo incendios (superficie ³1 ha) y conatos…
Después de un largo invierno y una triste primavera, ya está aquí el período estival y, como todos los años, empezamos a recibir las primeras noticias de incendios forestales que azotan nuestros bosques y destruyen nuestro entorno natural. El año pasado 2019 se contabilizaron alrededor de 10,883 incendios incluyendo incendios (superficie ³1 ha) y conatos de incendio (superficie <1 ha) constituyendo un total de 26,252 ha de superficie arbolada y 83,962 ha de superficie forestal. Los incendios, aparte de afectar de forma importante e irreversible nuestro patrimonio paisajístico, arrasan viviendas, e incluso poblaciones enteras en ocasiones provocando la muerte de sus habitantes y de miembros de los equipos de salvamento y extinción de incendios. No obstante, a pesar de la capacidad destructiva innegable de un incendio forestal, es posible la aparición de nueva vida de la devastación ya que existen numerosas especies vegetales que aprovechan dichos eventos, sean provocados o fortuitos, para colonizar nuevos ambientes y proliferar aprovechando que sus competidores han sido barridos por el incendio. Además, estas especies vegetales son capaces de aprovechar el humo procedente de la combustión de materia vegetal como señal, como los indios nativos americanos, aunque, en este caso no para avisar a sus compañeros sino para inducir respuestas de crecimiento y desarrollo que permitan una rápida colonización del terreno arrasado por las llamas.
En concreto, las plantas son capaces de reconocer distintos compuestos químicos procedentes de la combustión de materia vegetal que, a su vez, disparan una serie de mecanismos moleculares para inducir la germinación y proliferar más rápidamente que sus competidoras. Estas señales, identificadas en 19941, se llaman karrikinas (karrikins en inglés) (Figura 1) y derivan de la combustión de carbohidratos, azúcares y polisacáridos como la celulosa presente en las paredes de las células vegetales que provoca su conversión a piranobutenólidos, estructuralmente muy similares a otros compuestos fitoreguladores llamados estrigolactonas2, de los que ya hablaremos en otra ocasión.
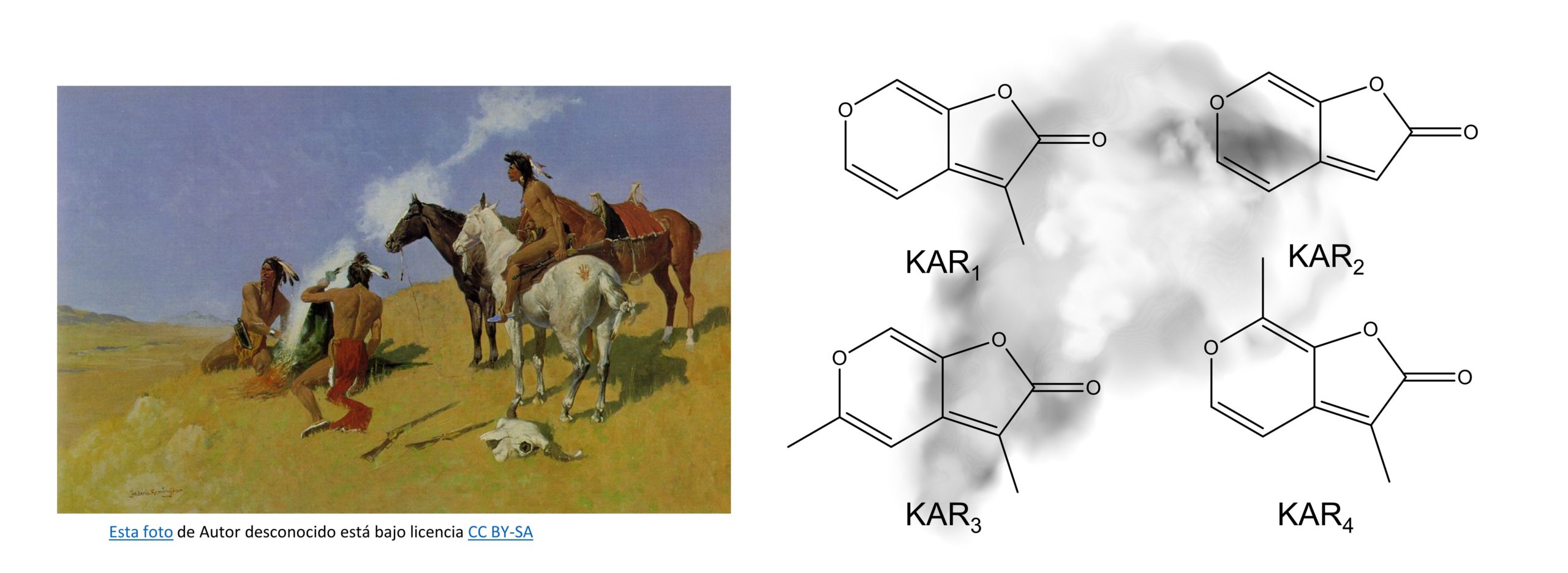
Indios nativos americanos y estructura de las diferentes karrikinas identificadas en el humo procedente de la combustión de material vegetal.
¿Cómo funciona este mecanismo? Bueno, para ello, tenemos que entender primero qué es una semilla y cómo las plantas desarrollan las semillas. Las semillas son órganos especializados en la dispersión y reproducción de las especies de vegetales superiores y, generalmente, contienen un embrión obtenido por fecundación de un óvulo (sí, ¡¡igual que nosotros!!), y una serie de tejidos que protegen y aportan nutrientes durante el proceso de germinación. Tras la fecundación, el óvulo madura y entra en un estado de quiescencia llamado dormición que evita que germinen antes de tiempo y que, en algunas especies vegetales, puede prolongarse cientos e incluso miles de años. Esta dormición asegura que los embriones germinarán y proliferarán solo cuando se den ciertas condiciones óptimas o se perciba determinado estímulo. En este sentido, y especialmente durante los procesos de inducción de la dormición y la germinación, las hormonas vegetales juegan un papel clave. Por ejemplo, durante la ontogenia de la semilla, ésta ha de adoptar una serie de características fisiológicas que le permitan aguantar períodos de sequía y otras adversidades y seguir siendo viable, en la regulación de este proceso la hormona ácido abscísico es fundamental. En cambio, la germinación, que requiere para su inicio unas condiciones de iluminación, temperatura y humedad, etc… determinadas (cada especie tienes unos requerimientos particulares), sigue el camino inverso: se reactiva la actividad metabólica del embrión, se sintetizan enzimas que liberan los nutrientes de los tejidos de reserva y que así estén disponibles para el crecimiento de la plantulita, etc y aquí participan otras hormonas, como por ejemplo las giberelinas…En este sentido, las semillas de especies cuya expansión está asociada a los períodos entre incendios también entran en dormición para posponer su germinación hasta que se alcancen las condiciones óptimas tras el incendio y, en su caso, que se perciban los estímulos físicos y químicos adecuados, como en este caso las karrikinas, entre otros factores2.
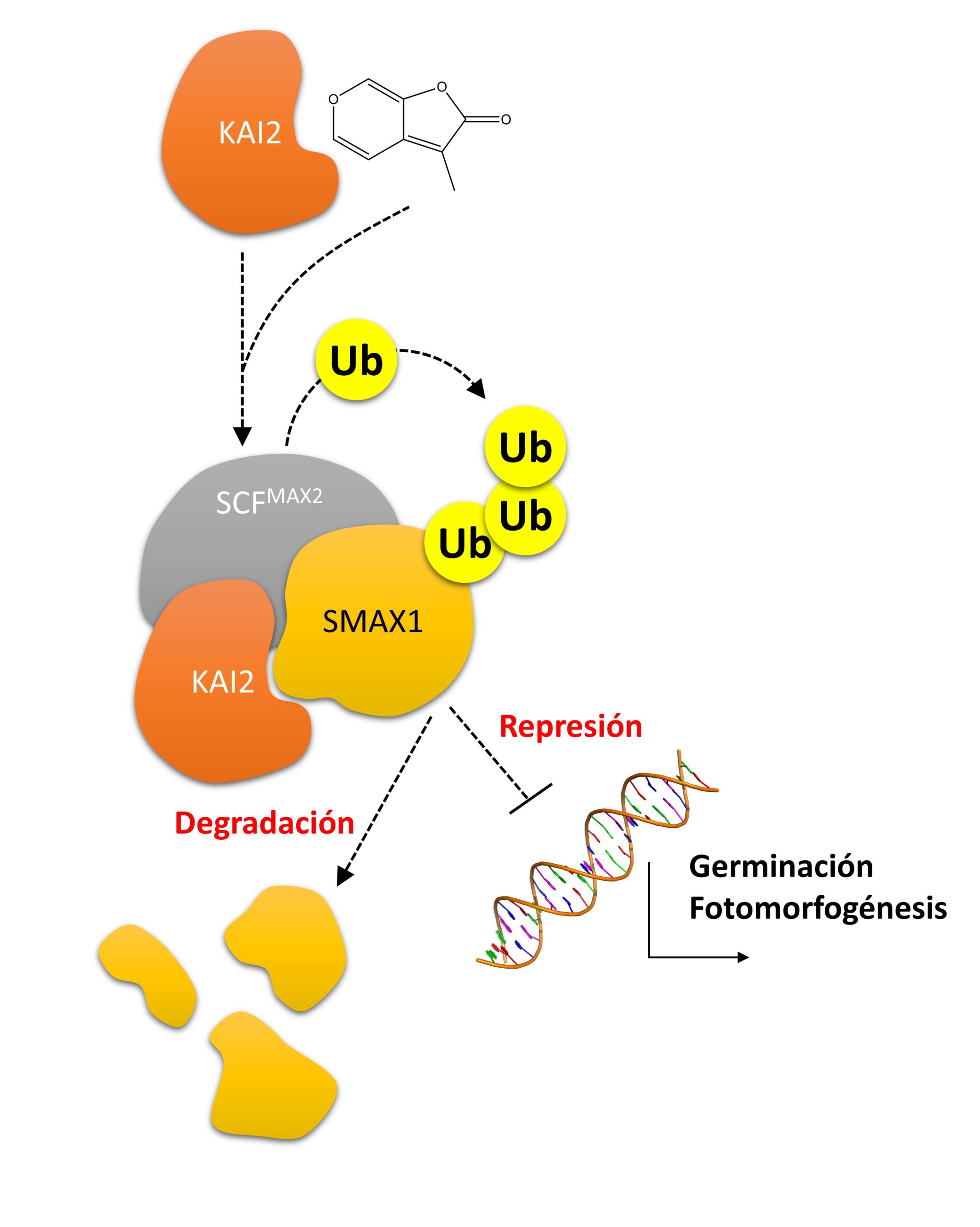
Esquema del mecanismo de señalización mediado por karrikinas. KAI2 es el receptor putativo de las karrikinas, SMAX1 es el represor de la expresión génica, SCFMAX2 es la proteína encargada de marcar SMAX1 para su degradación, Ub es la ubiqüitina, una pequeña proteína de marcaje. La unión de KAI2 promueve la formación del complejo SCFMAX2 y SMAX1, su marcaje con Ub y su degradación, permitiendo la expresión de los genes que estaba reprimiendo.
En todo proceso de señalización molecular, se necesita, aparte de la molécula señalizadora, un receptor, una proteína (o varías) que se unan a la molécula señalizadora y cuya unión provoque un cambio estructural o químico que actúe como relé para activar/desactivar el siguiente paso. Este proceso se llama percepción. En el caso de las karrikinas, se ha descrito que una proteína llamada KAI2 podría ser el receptor para estas moléculas cuya unión provocaría la degradación de proteínas represoras de la transcripción (SMAX1) mediante un mecanismo dependiente de ubiquitina (Ub), una pequeña proteína que sirve para marcar a otras para su degradación (Figura 2). La degradación de este represor SMAX1 permitiría la expresión de numerosos genes relacionados con la germinación, el desarrollo y elongación de brotes3. Se han identificado parientes lejanos de esta proteína KAI2 en eubacterias y dentro de los vegetales existen ortólogos en helechos, musgos, hepáticas y algas clorofíceas3. Por tanto, es una proteína con cierto pedigree, sugiriendo que el mecanismo que regula es de gran importancia, de ahí que se haya conservado tan bien durante la evolución de las plantas superiores. Los estudios sugieren que la especialización de esta proteína para la percepción de karrikinas pudo haber ocurrido durante el período cretácico (hace entre 65 y 126 Ma) cuando vastos bosques de coníferas cubrían gran parte de la superficie del planeta y las condiciones ambientales eran propicias para la ocurrencia de incendios. De hecho, las adaptaciones pirofíticas tales como el engrosamiento de la corteza del tronco, la serotinia o la dispersión de semillas estimulada por el fuego o la estimulación de la germinación por componentes del humo se estima que se originaron hace alrededor de 120 Ma, situando el fuego como una importante fuente de presión selectiva a mediados-finales del cretácico entre las plantas con flores2.
De las especies vegetales que presentan este tipo de adaptaciones, el género Pinus es posiblemente uno de los más importantes por su valor forestal y por el gran número de especies existentes (alrededor de 110 especies pertenecientes al género Pinus). Desde hace tiempo se conoce la extraordinaria capacidad de regeneración de los pinares tras un incendio, aunque actualmente no se considere una especie pirófita estricta ya que las elevadas temperaturas no favorecen la germinación de sus semillas, como pasa en otras especies de los géneros Cistus, Cytisus o Genista4. No obstante, y como ya se ha indicado antes, presentan importantes adaptaciones para vivir en hábitats susceptibles de incendiarse como la presencia de estructuras aislantes que protegen meristemos capaces de rebrotar tras el incendio (brotación epicórmica) o los conos (piñas) que protegen a las semillas del intenso calor y además facilitan su dispersión (serotinia), permitiendo que una importante fracción de ellas pueda germinar en los espacios abiertos generados tras el incendio5. Como se ha comentado anteriormente, las semillas de pino no responden bien al tratamiento exógeno con karrikinas, no pudiendo pues considerarse una especie pirófita. En contraposición, las especies pirófugas son ciertamente insensibles y no muestran un incremento en la capacidad de germinación tras el tratamiento con karrikinas6.
Todo esto demuestra, una vez más, que las plantas son organismos maravillosos capaces de adaptarse a las situaciones más diversas y adversas imaginables y que su carácter sésil no ha de considerarse una limitación sino una oportunidad para desarrollar otras formas de interaccionar con su entorno.
Este artículo nos lo envía Vicent Arbona. Doctor en Ciencias Biológicas y Profesor Titular en la Universitat Jaume I (Castelló de la Plana). Investiga en el papel de la regulación hormonal de las respuestas fisiológicas y metabólicas al ambiente en plantas, centrándose en la reconfiguración del metabolismo primario y secundario en condiciones en las que el oxígeno o los carbohidratos son limitantes. Si no fuera científico, sería artista, que es la segunda mejor manera de acercarse a la naturaleza. Twitter: @arbona_vicent
Referencias científicas y más información:
- Nelson, D. C., Flematti, G. R., Ghisalberti, E. L., Dixon, K. W. & Smith, S. M. Regulation of Seed Germination and Seedling Growth by Chemical Signals from Burning Vegetation. Annual Review of Plant Biology 63, 107–130 (2012).
- Waters, M. T., Scaffidi, A., Flematti, G. R. & Smith, S. M. The origins and mechanisms of karrikin signalling. Current Opinion in Plant Biology 16, 667–673 (2013).
- Morffy, N., Faure, L. & Nelson, D. C. Smoke and Hormone Mirrors: Action and Evolution of Karrikin and Strigolactone Signaling. Trends in Genetics 32, 176–188 (2016).
- Martinez-Sanchez, J., Marín, A., Herranz, J., Ferrandis, P. & De las Heras, J. Effects of high temperatures on germination of Pinus halepensis Mill. and P. pinaster Aiton subsp, pinaster seeds in southeast Spain. Vegetatio 24, 69–72 (2018).
- Escudero, A., Sanz, M. V., Pita, J. M. & Pérez-García, F. Probability of germination after heat treatment of native Spanish pines. Annals of Forest Science 56, 511–520 (1999).
- Leonard, J., West, A. G. & Ojeda, F. Differences in germination response to smoke and temperature cues in ‘pyrophyte’ and ‘pyrofuge’ forms of Erica coccinea (Ericaceae). International Journal of Wildland Fire 27, 562–568 (2018).





