

Los sistemas CRISPR-Cas constituyen el sistema inmune adaptativo de bacterias y arqueas, pero su relevancia durante años ha radicado en la capacidad de utilizarlos como nuevas herramientas de edición genética. Estos sistemas fueron descubiertos a finales de los 90 y principios del 2000 por Francisco Mojica, estudiando secuencias repetidas existentes en el genoma de arqueas…

Las herramientas de edición genética CRISPR cortan el ADN en lugares precisos del genoma. Fotografía: Lluís Montoliu
Los sistemas CRISPR-Cas constituyen el sistema inmune adaptativo de bacterias y arqueas, pero su relevancia durante años ha radicado en la capacidad de utilizarlos como nuevas herramientas de edición genética.
Estos sistemas fueron descubiertos a finales de los 90 y principios del 2000 por Francisco Mojica, estudiando secuencias repetidas existentes en el genoma de arqueas de las salinas de Santa Pola (Alicante) (ref 1-2), de aquí lo importante de invertir en investigación fundamental. Fue posteriormente en 2012 cuando investigadores de UC Berkeley (California, USA) y del Instituto Max Planck (Alemania), entendieron que esta(s) proteína(s), capaces de unirse al ADN y cortarlo, podían ser dirigidas mediante una guía de ARN indicando la localización exacta donde el ADN debe ser cortado (ref 3), desarrollando así toda una tecnología de edición genética basada en la capacidad para cortar el ADN de forma precisa y eficaz, y durante la reparación del corte, introducir la modificación deseada. La tecnología CRISPR es ya una realidad que permite hacer casi cualquier modificación genética en cualquier organismo vivo, desde bacterias hasta células humanas pasando por animales y plantas, desde el desarrollo de soluciones para enfermedades genéticas, inmunoterapia, nuevos antimicrobianos, hasta herramientas de diagnóstico e incluso desarrollo de nuevos alimentos.
Esta tecnología junto con sus aplicaciones en terapias y productos reales dio lugar a una serie de patentes de valor incalculable, una guerra abierta entre las más poderosas instituciones por los derechos de patentes (ref 4) y un gran desarrollo industrial con decenas de empresas aplicándola en distintos campos con un negocio multibillonario que ya es una realidad. Pero de esto hablaremos en otro texto.

La carrera imparable por el desarrollo tanto de nuevas aplicaciones como de nuevas tecnologías ha dado lugar al descubrimiento de una nueva generación de herramientas de edición genética. En 2018, investigadores del Instituto Nacional de Salud de USA (NIH) descubrían toda una nueva variante de sistemas CRISPR donde se detectaba la presencia de transposones asociados a CRISPR, y potencialmente ligados a su funcionalidad (ref 5). Hasta ahora nunca se había descrito nada similar.
Los transposones, descubiertos por Barbara McClintock en los 80 y que le supondría el Premio Nobel (ref 6), son secuencias de ADN móviles con autonomía propia para saltar o desplazarse de una región a otra del genoma, generalmente arrastrando una carga genética con ellos. Por su capacidad móvil, los transposones se han usado históricamente para generar mutaciones aleatorias a lo largo del genoma, interrumpiendo genes y estudiando su funcionalidad. Pero como decimos, era mutagénesis aleatoria ya que no se podía controlar el evento de transposición es decir, donde se iba a insertar el transposón ni la carga asociada.
La existencia de transposones tipo TniQ y Tn-7 ligados a sistemas CRISPR-Cas, tanto tipo I como tipo V, descritos en los trabajos de Eugene Koonin en 2018, supone un cambio completo de paradigma. Estos investigadores hipotetizaban que quizá a la capacidad móvil de los tranposones ahora se añadía la capacidad de dirigir donde estos actúan, usando la funcionalidad de CRISPR ya que estaban ligados en un sistema único, aunque todo era predictivo y descriptivo basado en análisis bionformáticos. Además, existe otra particularidad, en estos sistemas de transposones asociados a sistemas CRISPR-Cas, no existe una nucleasa capaz de cortar el ADN, como en un sistema CRISPR normal, por lo tanto, el mecanismo de actuación, tamaño y complejidad es distinto.
Sólo un año después, en Julio de 2019, se publicaban dos trabajos en Nature y Science (ref 7-8), donde se demostraba que efectivamente los transposon-encoded CRISPR-Cas systems podían utilizarse para edición genética y más en concreto, para realizar inserciones en el genoma.
En el caso del trabajo dirigido por Sam Sternberg, Universidad de Columbia, usaron el sistema de transposon Tn6677 asociado a CRISPR I-F identificado en la bacteria Vibrio cholera, y expresando todos sus componentes (genes TniQ-Cas8-Cas7-Cas6, genes TnsA-TnsB-TnsC, guia ARN, mas la secuencia a insertar) en Escherichia coli, demostraron que se podían llevar a cabo inserciones en el genoma de forma específica y fiable. Este sistema emplea la guía de ARN del sistema CRISPR (que indica a que secuencia de ADN unirse) y las proteínas Cas678 para formar un complejo de unión al ADN y reclutar los componentes del transposon TniQ-TnsABC para llevar a cabo el evento de transposición, insertando la secuencia de ADN deseada en el sitio deseado. Sus autores realizaron varias mutaciones en el genoma de E. coli, en distintos genes y con inserciones de distintos tamaños, para demostrar la versatilidad de la tecnología.
En el caso del trabajo de Feng Zhang, de BROAD-MIT, usaban el tranposon Tn7 asociado al sistema CRISPR V-K identificado en la cianobacteria Scytonema hofmanni, y como en el caso anterior, expresando todos sus componentes (TniQ-TnsB-TnsC, Cas12K, guía ARNA, y la secuencia a insertar) en E. coli demostraban la capacidad de realizar inserciones en el cromosoma.
En ambos casos hay dos particularidades que diferencian los sistemas de transposones asociados a CRISPR de los sistemas CRISPR normales, más allá de la existencia del transposón per se. La primera particularidad, es que en los sistemas mediados por transposones no existe una nucleasa Cas que corte el ADN. En Tn6677, asociado a CRISPR I-F, Cas3 que es la nucleasa tipo de sistemas CRISPR tipo I no está presente. En el sistema Tn7 asociado a Cas12K, tampoco existe nucleasa que corte el ADN ya que Cas12k no tiene capacidad catalítica para cortar. La ausencia de nucleasas, diferencia más significativa frente a un sistema estándar CRISPR-Cas, implica que la inserción de ADN es mediada por el evento de transposición ejecutado por el transposón, y no basado en cortar el ADN y repararlo introduciendo las modificaciones deseadas, como ocurre con CRISPR. La segunda diferencia, es que las inserciones en el genoma mediadas por los transposones asociados a CRISPR, ocurren a una distancia fija de la región de ADN indicada por la guía de ARN, ya sea de 40 ó 60 pares de bases dependiendo del sistema utilizado. Esto es un factor muy importante a la hora de tener en cuenta donde se va a llevar a cabo la inserción deseada y cómo hacer el diseño experimental.
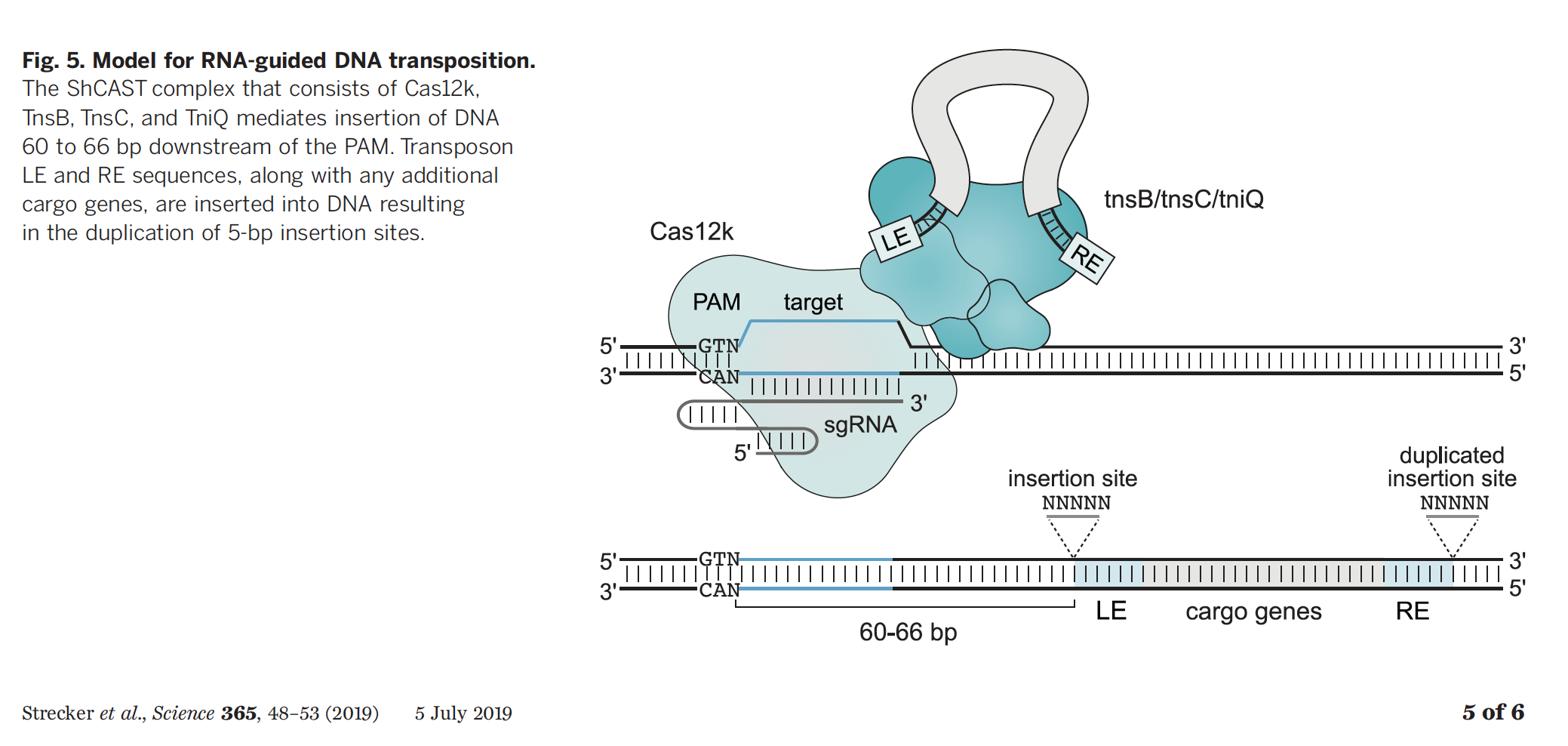
De nuevo S. Sternberg, publicaba a finales de 2019 una caracterización de la estructura tridimensional de Tn6677 para entender la interacción del complejo de proteínas Cas678 – TniQ-TnsABC, para mayor entendimiento del funcionamiento del transposon y su interacción con los componentes CRISPR (ref 9). Y ahora en 2020, el mismo grupo publicaba un nuevo avance y una mejora en todos los aspectos de la tecnología (ref 10). Todos los componentes necesarios de transposon Tn667 eran expresados en un único vector plasmídico (INTEGRATE), reduciendo así la complejidad del sistema y aumentando la eficiencia de la tecnología. Tal es la mejora que han llegado a hacer inserciones de hasta 10.000 nucleótidos (10Kb) con eficiencias del 100%. Además, combinando varias guías de ARN conseguían realizar varias inserciones genéticas de una sola vez. Más aún, combinando esta tecnología con integrasas y recombinasas conseguían por primera vez hacer otras modificaciones en el ADN como delecciones (eliminar), previa inserción de secuencias flanqueantes, constituyendo un sistema en dos pasos consecutivos. Todas estas optimizaciones además se han llevado a cabo en otras bacterias de interés médico e industrial como Klebsiella oxytoca y Pseudomonas putida respectivamente, y no sólo en E. coli.
Estos trabajos suponen la base del desarrollo de una nueva tecnología de edición genética programable, precisa y eficaz, que permite editar el ADN sin necesidad de cortarlo ni requiere de los mecanismos de reparación de ADN celular (principalmente recombinación homóloga) para introducir el cambio deseado durante la reparación, como ocurre con CRISPR. Aunque el desarrollo tecnológico no ha hecho más que empezar y todavía falta su validación en células Eucariotas, abre un nuevo camino aumentando así las herramientas disponibles para editar el ADN en todo tipo de aplicaciones y posiblemente en casi cualquier organismo. Además, esta nueva tecnología supone el desarrollo de nuevas patentes y derechos de explotación ofreciendo alternativas a la situación actual de CRISPR, lo cual ya se ha hecho notar con la creación de la primera empresa, Tessera Therapeutics, que explotará esta tecnología para aplicaciones terapéuticas en salud humana.
AUTOR: Este artículo nos lo envía Claudio Hidalgo Cantabrana, es Doctor en Biología por la Universidad de Oviedo y realizó su doctorado en microbiología y genética en el Instituto de Productos Lácteos de Asturias (IPLA-CSIC) en el ámbito de probióticos, microbiota y salud humana. Actualmente se encuentra trabajando como investigador en North Carolina State University (Raleigh, NC, USA) desarrollando nuevas herramientas de edición genética basadas en CRISPR-Cas para llevar a cabo ingeniería metabólica en bacterias. Durante su carrera profesional ha realizado estancias en distintos centros europeos, recibido premios y becas nacionales y europeas, es autor de una treintena de artículos SCI, con ponencias en congresos nacionales e internacionales. Ha desarrollado y obtenido varias patentes, es co-fundador y miembro del comité científico asesor de Microviable Therapeutics y co-fundador de CRISPR Biotechnologies Inc. Podéis seguirle en Twitter (@ClaudioCRISPR) o en su cuenta de Linkedin.
Referencias científicas y más información:
- https://pubmed.ncbi.nlm.nih.gov/7476211/
- https://pubmed.ncbi.nlm.nih.gov/15791728/
- https://pubmed.ncbi.nlm.nih.gov/22745249/
- https://naukas.com/2018/05/01/la-obviedad-la-disputa-las-patentes-crispr/
- https://pubmed.ncbi.nlm.nih.gov/29784811/
- https://www.nobelprize.org/prizes/medicine/1983/mcclintock/biographical/
- https://www.nature.com/articles/s41586-019-1323-z
- https://science.sciencemag.org/content/365/6448/48.long
- https://www.nature.com/articles/s41586-019-1849-0
- https://www.biorxiv.org/content/10.1101/2020.07.17.209452v1





